
9 Физиологиеские эффекты ДХМ
В этом разделе рассказывается о физиологических эффектах ДХМ в свете последних исследований о нем. ДХМ не очень хорошо изучен в
качестве рекреационного наркотика, многое из здесь представленного - предположения, некоторые - полностью мои. Чтобы быть точным,
многое из изложенного представляет собой "научно-гипотетическое пиздобольство" и должно восприниматьчся как интересные и
недоказанные гипотезы.
Большая часть этой главы очень техническая. В следующей (Раздел 10) вы найдете введение в нейрофармакологию; глоссарий (Раздел 16) содержит определения и объяснения технических терминов.
Это сложная проблема. Кашлевой рефлекс обычно включает в себя серии сигналов, исходящих из горла, легких, носоглотки и заканчивающихся в мускулах. В любой точке этого пути сигналы могут быть заблокированы. Сигма-рецепторы очевидно вовлечены в этот путь (42,49,55,56). Это может быть прямым влиянием - сигма-активация может напрямую подавлять сигналы кашлевого рефлекса - или непрямым. Эффект подавления кашля опиатами (типа кодеина) не сходен с тем же самым эффектом не-опиатных морфинов типа ДХМ (49); вместо этого, он управляeтся традиционными опиатными рецепторами (мю, каппа, дельта).
Есть факты, свидетельствующие о том, что 5HT1A-рецепторы (рецепторы серотонинового типа) также участвуют в передаче сигналов, и что подавители кашля могут увеличить 5HT1A активность (57), вероятно посредством NMDA-антагонизма (90). Это может объяснить некоторое изменяющее настроение воздействие ДХМ. 5HT1A-рецепторы работают в состоянии беспокойства и при сопротивлении неприятным событиям. Буспирон, частичный агонист 5HT1A-рецептора, транквилизатор менее сильный (но более безвредный), чем бензодиазепины типа диазепама (ValiumTM).
Если вы думаете, что кашлевой рефлекс сложный, то погодите!
ДХМ связывается покрайней мере с четырьмя местами связывания (сайтами) в мозге (58), которые могут быть произвольно помечены DM1, DM2, DM3, и DM4; вероятно существует и пятый сайт (DM5). Некоторые из этих сайтов восприимчивы к пентазоцину, известному сигма-лиганду; некоторые - к галоперидолу, другому сигма-лиганду. В следующей таблице упорядочена информация из нескольких источников. Приведено сродство к связыванию ДХМ, DTG и 3-PPP (58) вместе с восприимчивостью к (+)-пентазоцину (60) и способностью к замещению галоперидола (58) (связывание измеряетмя в нМ [наноМолях], кроме указанных случаев). "Низкое" означает микромолярное сродство к связыванию.
Эта таблица демонстрирует, что ДХМ связывается с четырмя различными сайтами, к двум из них с высоким сродством. Принято считать первый из них сигма1-рецептором, основываясь на связывании с пентазоцином и галоперидолом и силе действия (+)-3-PPP. Второй - почти наверняка PCP2-рецептор, принимая во внимание невосприимчивость к пентазоцину и очень сильное сродство с ДХМ. Третий - вероятно сигма2 (принимая во внимание влияние DTG), но вероятно что "DM1" в этой таблице представляет и сигма1 и сигма2, а третий сайт - что-то еще. Четвертый сайт, вероятно, являеется сайтом открытого канала NMDA-рецептора, хотя не исключено, что это место связывания ионного канала (59).
К сожалению, у меня нет информации о связывании ДХО (декстрорфана). Он вероятно проявит более сильное связывание в сайте DM4 (сайт открытого канала NMDA), затем с DM1 (сигма1), а после с DM3 (сигма2) и/или DM2 (PCP2), или обоими. В настоящее время я ищу эту информацию.
Сайт связывания PCP2 - это, вероятно, структура обратного всасывания дофамина, поэтому его блокада предотвратит поглощение дофамина путем, сходным с антидепрессантом бупропионом (WellbutrinTM) или кокаином. Разумеется, ДХМ существенно слабее кокаина (и сильнее бупропиона, между прочим) в этом сайте. Это вероятная причина эйфорических эффектов низкой рекреационной дозы, и почти наверняка объясняет стимулирующие эффекты малой дозы. Интересно, что стимулирующий эффект кажется качественно отличным от амфетаминов большинству людей (сравнительной информации для кокаина у меня нет). Один потребитель сравнил ДХМ и бупропион по благоприятному стимулирующему эффекту. Между прочим, возможно, что ДХМ неконкуррентно связывается с сайтом погложения дофамина, где бупропион связывается конкурирентно.
Музыкальная и двигательная эйфория частично вызваны PCP2 активностью. И NMDA блокада и сигма активность могут приводить к дофаминергической активности (см. ниже), торможение обратного поглощения усилит эти эффекты.
Интересно, что ДХМ вроде бы более сильно воздействует на этот сайт, чем другие сигма/NMDA лиганды (такие как PCP или кетамин) по сравнению с активностью в других сайтах. Также вызывает интерес, что по крайней мере один трициклический антидепрессант активен в родственных рецепторах (сигма, PCP) (71,74,75); возможно, что на сайт PCP2 могут быть нецелены некоторые антидепрессанты.
Так как сигма2 сайт довольно новое открытие, не известно какие сигма-подобные эффекты и поведение какому рецептору (сигма1 или сигма2) принадлежат. Очень мало данных о субъективных эффектах сигма-лигандов, частично потому, что только недавно стали доступны селективные лиганды, частично из-за нежелания исследователей это выяснить на самих на себе. ДХМ связывается с сигма1-рецептором и в основном полагается агонистом этого рецептора. Вероятно, что и в сигма2-рецепторе ДХМ является агонистом, хотя тут он более слаб.
Нарушение сенсорного процессинга частично может происходить из-за сигма-активации (и частично из-за NMDA блокады) (63-65). Сигма-рецепторы могут специфично вовлекаться в слуховые эффекты ДХМ (65), а эти эффекты могут иметь связь с разрушением устойчивого сенсорного процессинга.
Психотомиметические (психозоподобные) эффекты ДХМ могут быть результатом сигма активности, поскольку сигма-рецепторы вероятно каким-то образом вовлечены в шизофрению (46-49). Люди, применявшие и ДХМ и кетамин, отметили, что ДХМ более склонен к вызыванию бредовых и гиперабстрактных мыслешаблонов. Сигма-рецепторы могут временно модулировать холинергические рецепторы (97), поэтому сигма-активность может производить временные эффекты немного сходные с антихолинергиками, вызывающими бред. [что-то типа циклодола?]
Влияние на моторные навыки может оказаться результатом активности именно сигма2-рецепторов (69). как только сигма2-рецепторы будут изучены более полно, будет больше информации. Разумеется, это может быть и вкладом NMDA-рецепторов.
Декстрорфан (ДХО) более активен, чем ДХМ, в NMDA-рецепторе, что объясняет ме-е-е-едленное наступление эффектов ДХМ даже в случае инъекции. NMDA-рецептор - это центральное место действия диссоциатива, и возможно он дает основной вклад в эффекты ДХМ. Диссоциативы, включая ДХМ, действуют на NMDA-рецепторы, связываясь с его каналом, как тоько он откроется, плотно затыкая его.
Большинство "опьяняюших" эффектов, или интоксикация, ДХМ вызывается блокадой NMDA-рецептора. Опьяняющий эффект алкоголя вероятно частично осуществляется блокадой NMDA-рецепторов (депрессивный эффект алклголя происходит из-за ГАМК-активности; ДХМ не проявляет активности в ГАМК-рецепторах) (28,61,62). Диссоциативная анестезия высоких доз ДХМ происходит скорее всего тоже из-за блокады NMDA-рецепторов (63).
Как говорилось выше, нарушение сенсорного процессинга, особенно при высоких дозах ДХМ, частично происходит из-за воздействия на NMDA и частично - на сигма-рецепторы (63-65). NMDA-блокада вероятно несет ответственность за основную, если не всю, часть флэнджер-эффектов ДХМ, особенно визуального флэнджера. Скорее всего это непрямой эффект, т.е. блокада NMDA-рецепторов вызывает изменения в способе обработки мозгом информации, приводящие к флэнджеру.
Воздействие на память наверняка происходит из-за NMDA-блокады. NMDA-рецепторы глубоко вовлечены в договременную потенциацию (64,66-68) - первичный механизм перед промежуточной (кратковременной, рабочей) памятью (КВП) и долговременной (постоянной) памятью (ДВП). Блокирование NMDA-рецепторов вызывает нарушения долговременной потенциации, и следовательно кратковременной и постоянной памяти.
NMDA-блокада косвенно (244) увеличивает дофаминовую активность в полосатом теле, nucleus accumbens, обонятельных бугорках и префронтальной коре (204,226, 326). Увеличенная активность в дофаминовых D1-рецепторах приводит к усиленнию локомоторной активности, наблюдавшейся у крыс под воздействием диссоциативов (178), а также может нести ответственность за монгие другие воздействия ДХМ на движение. Хроническое потребление ДХМ может вызвать апрегуляцию (повышенную возбудимость) дофаминовых D2-рецепторов (205).
Способность ДХМ подавлять дыхание на токсических уровнях скорее всего результат блокады NMDA-рецепторов или (по моему мнению) блокады ионных каналов. Некоторые эффекты очень высоких доз могут происходить из-за всеобщего нарушения работы нейросетей. Предварительные факты свидетельствуют, что и эффект "самопроизвольных воспоминаний" и ощущения аналогичные опыту смерти (near death experience) могут происходить в тот момент, когда работа лимбических областей (гиппокамп, гиппокампальная формация и окружающие области) нарушена NMDA-блокадой. Большинство наркотиков нацеливаются на специфические, маленькие скопления (кластеры) нейронов (или их рецепторов, которые могут быть рассеяны вокруг). И вырабатывающие аминокислоты нейроны и NMDA-рецепторы имеют тенденцию к равномерному распределению, хотя и они сконцентрированы в определенных областях мозга, но очень плотно.
Вероятно наибольшее количество эффектов ДХМ проистекает из-за последствий его связывания с выщеуказанным рецептором в височных лимбических областях. Блокада NMDA-рецепторов ДХМ имеет специфические последствия в этих областях. Нормальное функционирование гиппокампа и миндалины мозжечка нарушается потому, что NMDA-блокада предотвращает долговременную потенциацию. В то же время, задняя часть коры поясной извилины и $retrosplenial cortex$ парадоксально активизируются. Возможны "шумовые" эффекты во всей лимбической системе, усиленные обратной связью. В одной статье предполагается, что диссоциативы могут вызывать "микроинсульты" или ограниченые очаги высокой активности в лимбической системе (176).
Предлагается и модель подавления ощущуний "сверху вниз", т.е. тормозящие импульсы из лимбической системы и окружающей корковой области посылаются "вниз" в сенсорные сети, уменьшая поток сенсорных данных. Такое торможение ощущуений "сверху вниз" может быть механизмом диссоциативной анестезии; сенсорная информация все еще обрабатывается мозгом, но никогда уже не проникает в сознание и не кодируется в промежуточной декларативной памяти (в гиппокампе).
Наконец, диссоциативы вероятно воздействуют на поток импульсов через лимбические области, возможно увеличивая степень внутренней обратной связи с этими зонами (или между этими зонами и неокортексом [новой корой]), затмевая тем самым входные сенсорные сигналы. Также возможно и изменение пропускания сигналов, связанных с тета-ритмом, через заднюю часть поясной извилины (311).
Сведение всего это вместе в некоторое подобие согласованной теории может быть преждевременным, но, в любом случае, я собираюсь проделать это. Я считаю (все это может перемениться с приходом новой информации), что пониженный уровень сенсорных данных (из-за торможения "сверну вниз"), ослабленное кодирование промежуточной памятью, сопряженное с расширением активности коры задней части поясной извилины и $retrosplenial cortex$, - все это приводит к усилению устойчивости петли обратной связи. В этой петле случайный "шум", индивидуальные различия в строении и "разводке" лобных долей, содержимое промежуточной памяти и влияние электромагнитных полей (332) в комбинации производят глубоко анормальнфе нейропаттерны.
Интересный побочный эффект всего этого - усиление способности распознавать запахи. Эта способность формально не изучалась, и на самом деле все может происходить только в голове, однако обонятельные сигналы обрабатываются мозгом немного по-другому, чем остальные, и они могут не попадать под тормозящие импульсы, ослабляющие другие сенсорнные сигналы. И если это так, то петля обратной связи может действительно служить усилителем обонятельных данных, постоянно прибавляя одни и теже маленькие сигналы друг к другу.
Многие из эффектов ДХМ несомненно возникают из-за его непрямой активности в других медиаторных системах. Например, он может ненапрямую увеличить 5HT [серотонин] активность, особенно в 5HT1A-рецепторе. Это может объяснить способность ДХМ изменять настроение. Еще один пример - дофаминергическая активность; ДХМ довольно сильно влияет на увеличение дофаминовой активности (и из-за активации сигма-рецепторов, и из-за предотвращения обратного захвата дофамина в PCP2-сайтах) (72,76). Так же показано, что блокада NMDA-рецепторов усиливает дофаминовую активность, а также активность других медиаторных систем (100).
Одним из выдающихся эффектов ДХМ - флэнджер входящих сенсорных сигналов. В разной мере это проявляется у различных наркотиков. Ниже я изложу свою гипотезу на эту тему. Заметим, в частности, связь флэнджера с торчем [stoning] и опьянением [buzzing] - в некотором роде флэнджер - это более глубокая степень торча. Некоторые отметили флэнджер или строб-эффект после выкуривания большого количества каннабиса, потребители закиси азота знакомы с флэнджером звуков. Этот эффект может производить и алкоголь.
У всех этих веществ одна общая черта - они способны тормозить долговременную потенциацию в гиппокампе. Есть предположения, что существует несколько наборов сигналов, проходящих через лимбические области. Сигналы в фазе с тета-ритмом и остальные проходят по-разному. Вероятно один набор сигналов сильнее вовлекаются в процесс запоминания, а остальные - сильнее в проведение сенсорных данных. Или, вероятно, что все наркотики, тормозящие долговременную потенциацию, активизируют кору задней части поясной извилины. В обоих случаях, прохождение сигналов в одной фазе c тета-ритммм может быть нарушено, что приведет к пульсирующему восприятию в темпе тета-ритма.
Есть и другая теория - нейросети в мозгу повторно обрабатывают одни и те же данные, пока не сформируется устойчивая кофигурация, а ДХМ (и другие наркотики) ослабляют и замедляют этот процесс. Когда одни нейросети заторможены, а другие работают с нормальной скоростью, характеристические частоты двух наборов сигналов будут различаться, приводя к "биению частоты" почти тем же самым путем, как и два очень близких звука приводят к биению частоты (краткая демонстрация этого - послушайте как настраивают гитару на слух).
Другой интересный эффект ДХМ - эго способность вызывать своеобразные когнитивные нарушения, которые я объединяю под термином "гиперабстракция". Два примера:
Многие вызванные ДХМ мысли включают в себя то, что иногда называется "Странными Циклами" в логике. Точно так же как и самопротиворечивое утверждение "это утверждение ложно", некоторые из них не могут быть воплощены в логической форме; другие - могут, но только в качестве гипотез. Мыслить на таком уровне абстракции очень нелегко (если вы конечно не Курт Гедль).
Некоторые люди, описавшие от первого лица случаи психоза и шизофрении, отметили увеличение абстрактных способов мышления (на ум приходит книга Дзен и Искусство Ухода За Мотоциклом Роберта М. Персига). Это может быть, разумеется, и полной чушью - эти абстрактные мыслеформы просто усиливающееся безумие (а в этом случае трудносопоставимы с конкретными идеями). С другой стороны, может быть, что нечто типа шизофрении и психотического состояния связано с "размазыванием" между уровнями абстракции. При достаточном "размазывании", мысль, немогущая быть представленной в деталях на конкретном уровне абстракции, сможет стать представимой в сознании.
Итак, ДХМ может вызвать нечто типа временного "размазывания" этих уровней абстракции. Происходит ли это благодаря NMDA или сигма-активности, я не знаю, хотя подозреваю последнюю, поскольку другие NMDA-антагонисты не имеют тенденции производить такие изменения в образе мышления.
Как говорилось выше, сигма-активность может модулировать холинергические рецепторы в мозге (97), что приводит к временному снижению их функционирования аналогично (но гораздо более безвредному) действию антихолинергиков - атропина, скополамина, циклизина (Marezine) и т.д. Известно, что холинергическая активность важна для запоминания, а многие ноотропы (Умные наркотики - "Smart Drugs") усиливают холинергическую функцию. Сигма-активность может на время понизить эффективность в некоторых холинергических рецепторах, что приводит к нарушению памяти и мыслительного процесса. Некоторые люди фактически отмечали, что ДХМ создает ощущение временной тупости (Тупые наркотики, а?), хотя это происходит не со всеми.
Многие биогенные аминовые системы, видимо, играют управляющую роль, и многие исследователи считают, что эти управляющие системы работают как "пусковые кнопки". Например, одна из теорий ЛСД говорит, что он изменяет "коэффициент усиления" распознающих нейросетей (вероятно уменьшая торможение, хотя и иными путями, чем постулировано для диссоциативов). Как следствие, случайный шум (необходимый для работы любой нейросети) становится гораздо сильнее входящей сенсорной информации. Сенсорное распознавание становится все менее и менее точным - следовательно - галлюцинации.
Эффекты ЛСД почти наверняка более сложны, чем описано, но идея "пусковых кнопок" может быть справедливой и в этом случае. Если это так, то холинергические системы могут быть "пусковыми кнопками" для когнитивных сетей в той же мере, какими 5HT2A/5HT2C-системы являются для сенсорных нейросетей распознавания. Бред может быть просто когнитивным эквивалентом галлюцинаций. Или другими словами, разница между ощущением себя "похожим на цветок" и мыслями "Я и ЕСТЬ цветок" может быть ответом на вопрос какая нейросеть поражена.
Проблемы с памятью в некотором роде происходят из-за NMDA-блокады, хотя некоторые потребители кетамина отмечали, что ДХМ сильнее влияет на память, чем кетамин. Возможно, что в добавок к наведению бредовых мыслей, ухудшение холинергических функций может быть виновно в проблемах с памятью. Это разумеется согласуется с эффектом галлюциногенных (delusional) антихолинергиков [как, например, тарен].
Между прочим, антихолинергики тоже воздействуют на ацетилхолиновые рецепторы, которые управляют работой сердца и дыханием (эти рецепторы видимо не управляются посредством сигма-активности). Рекреационное использование антихолинергиков может быть чрезвычайно опасным, приводя к коллапсу дыхания и сердечной недостаточности.
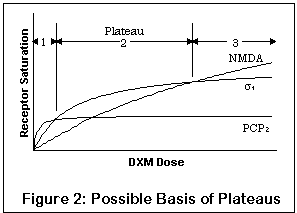 На рисунке изображен вероятный эффект совокупности рецепторов. Заметьте - это качественный рисунок, а не количественный;
настоящие уровени насыщения различных рецепторов для данной дозы ДХМ все еще практически неизвестны. Кривые означают уровни
насыщения в PCP2, сигма1 и сайте связывания в открытом канале NMDA, полное насыщение соответствующего
рецептора обозначено "уплощением" кривой. Ось Х -концентрация ДХМ; Y - процент насыщения рецепторов.
На рисунке изображен вероятный эффект совокупности рецепторов. Заметьте - это качественный рисунок, а не количественный;
настоящие уровени насыщения различных рецепторов для данной дозы ДХМ все еще практически неизвестны. Кривые означают уровни
насыщения в PCP2, сигма1 и сайте связывания в открытом канале NMDA, полное насыщение соответствующего
рецептора обозначено "уплощением" кривой. Ось Х -концентрация ДХМ; Y - процент насыщения рецепторов.
Между прочим, если вы знакомы с вещами подобного рода, вам может быть удобнее пользоваться координатами Lineweaver-Burk [что это - я не знаю], но для непосвященных это может оказаться смущающим.
Из-за повышенного сродства к PCP2, сигма1 и NMDA-рецепторам соответственно (сигма2 не представлен), малая доза будет оказывать пропорционально больший эффект на PCP2-рецепторы, в то время как увеличение дозы приведет к насыщению этих рецепторов. Прием большего количества ДХМ не изменит уровень насыщения PCP2 существенным образом, а окажет влияние на другие рецепторы.
Более того, более слабые эффекты воздействия на PCP2-рецепторы может быть почти полностью сглажены эффектами сигма1 и NMDA-рецепторов (различные уровни насыщения показывают это). Это совершенно разумно, поскольку и сигма1 и NMDA-активность выявляют довольно сильное изменение поведения, последний в большей степени, чем первый.
Итак, первое плато может соответствовать доминированию PCP2-активности с некоторой сигма-активностью и небольшим эффектом NMDA-блокады; второе плато - сигма и некоторым NMDA-эффектам; третье - глубокой NMDA-блокаде.
Это, несомненно, упрощение, и не принимает во внимание индивидуальные отклонения в рецепторах. Некоторые люди могут иметь генетические варианты рецепторов, к которым ДХМ имеет более сильное (или слабое) сродство. Человек с PCP2-рецепторами, сильно связывающимися с ДХМ, может наслаждаться стимулирующими эффектами гораздо больше других.
К тому же, ионные каналы и сигма2-рецепторы в рисунке не указаны. Однако они несомненно вносят свой вклад, и некоторые люди утверждают, что есть плато и между первыми тремя. Они могут быть неярко выраженными и незаметными для большинства.
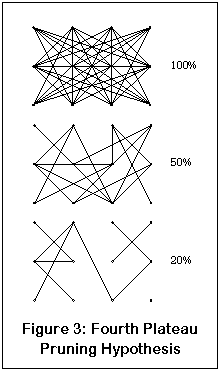
А что происходит на четвертом плато? Ну, вот еще один маленький рисуночек, который, надеюсь, прояснит все (ага, точно). Это рисунок нейросети; точки - нейроны, линии - их соединения. Как и все в мозгу, этоа нейросеть сильно связана. Проценты - количество оставшихся функциональных связей.
Когда достаточное число NMDA-рецепторов блокировано, нейрон может потерять большую часть входящей информации от другого нейрона и соединение эффективно нарушается. В начале это не причиняет неудобств, поскольку нейроны и связи нейронов еще достаточно плотны и другие нейроны могут взять на себя дополнительную нагрузку (хотя результатом может стать более медленная и менее аккуратная работа нейросети). В некоторый момент, достаточная связанность теряется и нейросеть уже больше не может функционировать.
Сравните это с диссоциацией четвертого плато. На некотором уровне, некие части мозга (вероятно гиппокамп) теряет способность к функцинированию настолько, что больше не может работать как единый блок. Сенсорный процессинг останавливается, и необработанная входящая сенсорная информация не может быть преобразована в соответствующе проанализированный (parsed) выходной поток. Сознание остается без реальной информации; вместо этого образуются хаотические нестабильные мыслеформы (паттерны). Следовательно - диссоциативная анестезия.
С другой стороны, может случиться так, что порог четвертого плато достигается, когда сенсорная информация тормозится достаточно для того, чтобы нейросигналы в лимбических областях отражались назад (feed back) на вход без существенного подавления со стороны входящей сенсорнной информации. Эта петля обратной связи продолжает возмущать нейропаттерны до тех пор, пока они не становятся оторванными от состояния человека или его воспоминаний.
ДХМ сам по себе очень сложное вещество; большинство наркотиков связыватся с одним или двумя рецепторами (покрайней мере с одним классом рецепторов). Рекреационное злоупотребление им, известное уже догие годы, еще не изучалось должным образом, он может воздействовать по-разному на разных людей. Рецепторы и сайты связывания, на которые он воздействует - сигма, NMDA и PCP2 - недавние открытия. Все это приводит к сложному и полохо понимаемому наркотику.
Кроме того, мозг - это сложная система, и, по большому счету, мы ничего не понимаем в его работе. Основы нейропередачи (нейротрансмиссии) вроде бы поняты, но остается много вопросов. Никто не знает почему так много нейромедиатров или подтипов рецепторов. Еще хорошо не изучены системы вторичных посредников большинства рецепторов. Большинство событий внутри нейронов происходит путем изменений в экспрессии генов, а это отдельная тема, о которой мало известно.
Повторю часто цитируемые (и правильные) слова: если бы наш мозг можно было бы просто понять, мы бы были такими примитивными, что не смогли бы его понять. Я верю, что в конце концов мы узнаем как работает мозг, но это случится не при моей жизни. До тех пор мы просто пытаемся догадаться, шаря в темноте.
ДХМ, как соль гидробромида, быстро абсорбируется из желудочно-кишечного тракта; в течении 30 минут он может войти в кроветок (2,3). Соединение полистирекса предназначено для продолжительной абсорбции, и его полный переход в кровь может занять 6-8 часов.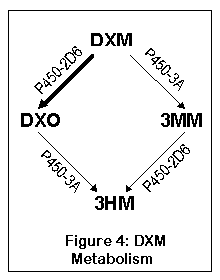
ДХМ метаболизируется двумя путями, оба из них приводят к одному - 3-гидроксиморфинану (3ГМ, 3HM). Первый путь исходит из ДХМ к ДХО (декстрорфан), а затем к 3HM; второй из ДХМ к 3-метоксиморфинану (3ММ), а затем к 3HM. Большая часть ДХМ (до 90%) метаболизуется через ДХО у нормальных индивидуумов.
ДХМ преобразуется в ДХО печеночным ферментом, называемым цитохром P450-2D6 (дебрисохин 4-гидроксилаза, debrisoquine 4-hydroxylase). Примерно у 7% кавказцев и 0,5% азиатов версия этого фермента работает крайне (до 70 раз медленнее) неэффективно и не может метаболизировать ДХМ в ДХО должным образом (10). От 0.5 до 2% популяции имеют многочисленные копии гена P450, что приводит к очень быстрому метаболизму ДХМ в ДХО (155). После превращения в ДХО, ферменты P450-3A4 и P450-3A5 перобразовывают ДХО в 3-гидроксиморфинан(77).
Другой путь ведет сначала к 3-метоксиморфинану (посредством ферментов P450-3A4 и P450-3A5), а уже затем к 3-гидроксиморфинану. Большинство людей не метаболизуют основное количество ДХМ таким образом, хотя люди с недостатком нормального P450-2D6 будут превращать существенную часть ДХМ в 3ММ. А 3ММ вероятно не психоактивен, что означает - 5-10% людей с недостатком фермента 2D6 ощутят меньший эффект от ДХМ (или более точно, не испытают эффектов ДХО).
P450-2D6 работает, убирая метокси группу и заменяя ее гидрооксилом (OH) (или точнее, отрезая метил от кислорода); этот процесс называется О-деметиляцией. P450-3A4 и 3A5 заменяют метильную группу водородом (H); это процесс N-деметиляции. Положение метильной и метокси групп показано на рисунке.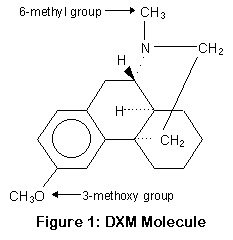
Как говорилось ранее, у некоторых людей отсутствует нормальный фермент P450-2D6. У остальной популяции этот фермент может быть ингибирован несколькими факторами. Многие лекарства ингибируют P450-2D6, особенно флюокситин (ProzacTM). Неполный список лекарств, ингибирующих фермент P450-2D6, дан в Разделе 15.1.
Сам по себе ДХМ, разумеется, будет конкурировать с другими веществами за P450-2D6 и, что важно, это будет делать и 3-метоксиморфинан (3ММ) (17). Фактически, 3ММ имеет большее сродство к ферменту P450-2D6, чем ДХМ. Это может объяснить тот факт, что вторая доза ДХМ "в догонку" призводит эффект отличный от первой. Конкуренция за фермент P450-2D6 уменьшит количество ДХМ, превращающегося в ДХО у второй дозы.
Этот график получен путем компьютерной симуляции метаболизма ДХМ: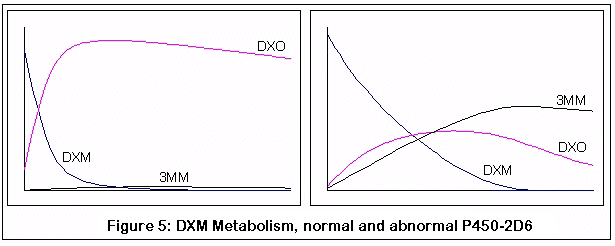
Первая пара графиков представляет метаболизм ДХМ у нормального индивидуума (слева) и индивидуума без нормального фермента P450-2D6 (справа). Заметьте быстрое и почти полное превращение ДХМ в ДХО у нормального индивидуума, по сравнению с менее эффективным и более медленным превращением у человека с недостатком P450-2D6.
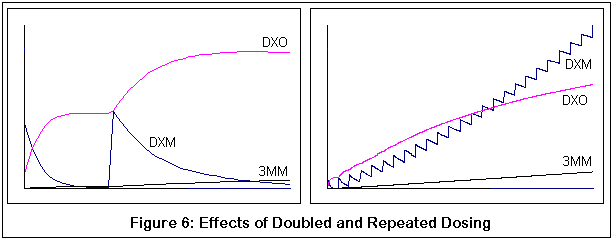
Вторая пара демонстрирует возможные результаты приема дополнительных доз ("в догонку"). Оба графика соответствуют нормальному варианту фермента P450-2D6. Отмечу, что вторая доза ДХМ не превращается в ДХО также быстро (поэтому меньший наклон). Правый график показыавет многочисленные дозы, и выгибание вверх метаболитической кривой ДХМ становится все более явным с каждой дозой.
Между прочим, это все качественные симуляции, а не количественные. Я пытался придерживаться известных значений КМ и VMAX для подходящих реакций [?], но симуляции были дискретным процессом (если честно, мои познания в уравнениях довольно слабы и требуют от меня титанических усилий). Я сравнил мои результаты с обнаруженными скудными данными, сравнения оказались разумными, но, опять же, я могу быть абсолютно неправым. Цель этих графиков - продемонстрировать относительные эффекты изменений в ферментной активности (из-за генетических изменений и конкурентного ингибирования 3-метоксиморфинаном), надеюсь, что этого оказалось достаточно.
У меня нет информации о том, что случается с самим 3-гидроксморфинаном. Он может выделяться напрямую почками, или подлежит дальнейшему метаболизму.
Большая часть этой главы очень техническая. В следующей (Раздел 10) вы найдете введение в нейрофармакологию; глоссарий (Раздел 16) содержит определения и объяснения технических терминов.
9.1 Каким образом ДХМ ингибирует кашлевой рефлекс?
Это сложная проблема. Кашлевой рефлекс обычно включает в себя серии сигналов, исходящих из горла, легких, носоглотки и заканчивающихся в мускулах. В любой точке этого пути сигналы могут быть заблокированы. Сигма-рецепторы очевидно вовлечены в этот путь (42,49,55,56). Это может быть прямым влиянием - сигма-активация может напрямую подавлять сигналы кашлевого рефлекса - или непрямым. Эффект подавления кашля опиатами (типа кодеина) не сходен с тем же самым эффектом не-опиатных морфинов типа ДХМ (49); вместо этого, он управляeтся традиционными опиатными рецепторами (мю, каппа, дельта).
Есть факты, свидетельствующие о том, что 5HT1A-рецепторы (рецепторы серотонинового типа) также участвуют в передаче сигналов, и что подавители кашля могут увеличить 5HT1A активность (57), вероятно посредством NMDA-антагонизма (90). Это может объяснить некоторое изменяющее настроение воздействие ДХМ. 5HT1A-рецепторы работают в состоянии беспокойства и при сопротивлении неприятным событиям. Буспирон, частичный агонист 5HT1A-рецептора, транквилизатор менее сильный (но более безвредный), чем бензодиазепины типа диазепама (ValiumTM).
9.2 Как ДХМ вызывает свои психоактивные эффекты?
Если вы думаете, что кашлевой рефлекс сложный, то погодите!
9.2.1 Общая информация
ДХМ связывается покрайней мере с четырьмя местами связывания (сайтами) в мозге (58), которые могут быть произвольно помечены DM1, DM2, DM3, и DM4; вероятно существует и пятый сайт (DM5). Некоторые из этих сайтов восприимчивы к пентазоцину, известному сигма-лиганду; некоторые - к галоперидолу, другому сигма-лиганду. В следующей таблице упорядочена информация из нескольких источников. Приведено сродство к связыванию ДХМ, DTG и 3-PPP (58) вместе с восприимчивостью к (+)-пентазоцину (60) и способностью к замещению галоперидола (58) (связывание измеряетмя в нМ [наноМолях], кроме указанных случаев). "Низкое" означает микромолярное сродство к связыванию.
| ДХМ сайт: | DM1 | DM2 | DM3 | DM4 |
|---|---|---|---|---|
| Вероятный сайт связывания: | сигма1 | PCP2 | сигма2 | NMDA |
| DXM | 50-83 | 8-19 | низкое | низкое |
| (+)-3-PPP | 24-36 | низкое | 210-320 | низкое |
| DTG | 22-24 | ??? | 13-16 | ??? |
| Восприимчив к пентазоцину? | да | нет | ??? | ??? |
| Замещает галоперидол? | нет | ??? | да | ??? |
Эта таблица демонстрирует, что ДХМ связывается с четырмя различными сайтами, к двум из них с высоким сродством. Принято считать первый из них сигма1-рецептором, основываясь на связывании с пентазоцином и галоперидолом и силе действия (+)-3-PPP. Второй - почти наверняка PCP2-рецептор, принимая во внимание невосприимчивость к пентазоцину и очень сильное сродство с ДХМ. Третий - вероятно сигма2 (принимая во внимание влияние DTG), но вероятно что "DM1" в этой таблице представляет и сигма1 и сигма2, а третий сайт - что-то еще. Четвертый сайт, вероятно, являеется сайтом открытого канала NMDA-рецептора, хотя не исключено, что это место связывания ионного канала (59).
К сожалению, у меня нет информации о связывании ДХО (декстрорфана). Он вероятно проявит более сильное связывание в сайте DM4 (сайт открытого канала NMDA), затем с DM1 (сигма1), а после с DM3 (сигма2) и/или DM2 (PCP2), или обоими. В настоящее время я ищу эту информацию.
9.2.2 Вклад сайта связывания PCP2
Сайт связывания PCP2 - это, вероятно, структура обратного всасывания дофамина, поэтому его блокада предотвратит поглощение дофамина путем, сходным с антидепрессантом бупропионом (WellbutrinTM) или кокаином. Разумеется, ДХМ существенно слабее кокаина (и сильнее бупропиона, между прочим) в этом сайте. Это вероятная причина эйфорических эффектов низкой рекреационной дозы, и почти наверняка объясняет стимулирующие эффекты малой дозы. Интересно, что стимулирующий эффект кажется качественно отличным от амфетаминов большинству людей (сравнительной информации для кокаина у меня нет). Один потребитель сравнил ДХМ и бупропион по благоприятному стимулирующему эффекту. Между прочим, возможно, что ДХМ неконкуррентно связывается с сайтом погложения дофамина, где бупропион связывается конкурирентно.
Музыкальная и двигательная эйфория частично вызваны PCP2 активностью. И NMDA блокада и сигма активность могут приводить к дофаминергической активности (см. ниже), торможение обратного поглощения усилит эти эффекты.
Интересно, что ДХМ вроде бы более сильно воздействует на этот сайт, чем другие сигма/NMDA лиганды (такие как PCP или кетамин) по сравнению с активностью в других сайтах. Также вызывает интерес, что по крайней мере один трициклический антидепрессант активен в родственных рецепторах (сигма, PCP) (71,74,75); возможно, что на сайт PCP2 могут быть нецелены некоторые антидепрессанты.
9.2.3 Вклад сигма сайта связывания
Так как сигма2 сайт довольно новое открытие, не известно какие сигма-подобные эффекты и поведение какому рецептору (сигма1 или сигма2) принадлежат. Очень мало данных о субъективных эффектах сигма-лигандов, частично потому, что только недавно стали доступны селективные лиганды, частично из-за нежелания исследователей это выяснить на самих на себе. ДХМ связывается с сигма1-рецептором и в основном полагается агонистом этого рецептора. Вероятно, что и в сигма2-рецепторе ДХМ является агонистом, хотя тут он более слаб.
Нарушение сенсорного процессинга частично может происходить из-за сигма-активации (и частично из-за NMDA блокады) (63-65). Сигма-рецепторы могут специфично вовлекаться в слуховые эффекты ДХМ (65), а эти эффекты могут иметь связь с разрушением устойчивого сенсорного процессинга.
Психотомиметические (психозоподобные) эффекты ДХМ могут быть результатом сигма активности, поскольку сигма-рецепторы вероятно каким-то образом вовлечены в шизофрению (46-49). Люди, применявшие и ДХМ и кетамин, отметили, что ДХМ более склонен к вызыванию бредовых и гиперабстрактных мыслешаблонов. Сигма-рецепторы могут временно модулировать холинергические рецепторы (97), поэтому сигма-активность может производить временные эффекты немного сходные с антихолинергиками, вызывающими бред. [что-то типа циклодола?]
Влияние на моторные навыки может оказаться результатом активности именно сигма2-рецепторов (69). как только сигма2-рецепторы будут изучены более полно, будет больше информации. Разумеется, это может быть и вкладом NMDA-рецепторов.
9.2.4 Вклад NMDA рецептора
Декстрорфан (ДХО) более активен, чем ДХМ, в NMDA-рецепторе, что объясняет ме-е-е-едленное наступление эффектов ДХМ даже в случае инъекции. NMDA-рецептор - это центральное место действия диссоциатива, и возможно он дает основной вклад в эффекты ДХМ. Диссоциативы, включая ДХМ, действуют на NMDA-рецепторы, связываясь с его каналом, как тоько он откроется, плотно затыкая его.
Большинство "опьяняюших" эффектов, или интоксикация, ДХМ вызывается блокадой NMDA-рецептора. Опьяняющий эффект алкоголя вероятно частично осуществляется блокадой NMDA-рецепторов (депрессивный эффект алклголя происходит из-за ГАМК-активности; ДХМ не проявляет активности в ГАМК-рецепторах) (28,61,62). Диссоциативная анестезия высоких доз ДХМ происходит скорее всего тоже из-за блокады NMDA-рецепторов (63).
Как говорилось выше, нарушение сенсорного процессинга, особенно при высоких дозах ДХМ, частично происходит из-за воздействия на NMDA и частично - на сигма-рецепторы (63-65). NMDA-блокада вероятно несет ответственность за основную, если не всю, часть флэнджер-эффектов ДХМ, особенно визуального флэнджера. Скорее всего это непрямой эффект, т.е. блокада NMDA-рецепторов вызывает изменения в способе обработки мозгом информации, приводящие к флэнджеру.
Воздействие на память наверняка происходит из-за NMDA-блокады. NMDA-рецепторы глубоко вовлечены в договременную потенциацию (64,66-68) - первичный механизм перед промежуточной (кратковременной, рабочей) памятью (КВП) и долговременной (постоянной) памятью (ДВП). Блокирование NMDA-рецепторов вызывает нарушения долговременной потенциации, и следовательно кратковременной и постоянной памяти.
NMDA-блокада косвенно (244) увеличивает дофаминовую активность в полосатом теле, nucleus accumbens, обонятельных бугорках и префронтальной коре (204,226, 326). Увеличенная активность в дофаминовых D1-рецепторах приводит к усиленнию локомоторной активности, наблюдавшейся у крыс под воздействием диссоциативов (178), а также может нести ответственность за монгие другие воздействия ДХМ на движение. Хроническое потребление ДХМ может вызвать апрегуляцию (повышенную возбудимость) дофаминовых D2-рецепторов (205).
Способность ДХМ подавлять дыхание на токсических уровнях скорее всего результат блокады NMDA-рецепторов или (по моему мнению) блокады ионных каналов. Некоторые эффекты очень высоких доз могут происходить из-за всеобщего нарушения работы нейросетей. Предварительные факты свидетельствуют, что и эффект "самопроизвольных воспоминаний" и ощущения аналогичные опыту смерти (near death experience) могут происходить в тот момент, когда работа лимбических областей (гиппокамп, гиппокампальная формация и окружающие области) нарушена NMDA-блокадой. Большинство наркотиков нацеливаются на специфические, маленькие скопления (кластеры) нейронов (или их рецепторов, которые могут быть рассеяны вокруг). И вырабатывающие аминокислоты нейроны и NMDA-рецепторы имеют тенденцию к равномерному распределению, хотя и они сконцентрированы в определенных областях мозга, но очень плотно.
9.2.5 Участие височных областей
Вероятно наибольшее количество эффектов ДХМ проистекает из-за последствий его связывания с выщеуказанным рецептором в височных лимбических областях. Блокада NMDA-рецепторов ДХМ имеет специфические последствия в этих областях. Нормальное функционирование гиппокампа и миндалины мозжечка нарушается потому, что NMDA-блокада предотвращает долговременную потенциацию. В то же время, задняя часть коры поясной извилины и $retrosplenial cortex$ парадоксально активизируются. Возможны "шумовые" эффекты во всей лимбической системе, усиленные обратной связью. В одной статье предполагается, что диссоциативы могут вызывать "микроинсульты" или ограниченые очаги высокой активности в лимбической системе (176).
Предлагается и модель подавления ощущуний "сверху вниз", т.е. тормозящие импульсы из лимбической системы и окружающей корковой области посылаются "вниз" в сенсорные сети, уменьшая поток сенсорных данных. Такое торможение ощущуений "сверху вниз" может быть механизмом диссоциативной анестезии; сенсорная информация все еще обрабатывается мозгом, но никогда уже не проникает в сознание и не кодируется в промежуточной декларативной памяти (в гиппокампе).
Наконец, диссоциативы вероятно воздействуют на поток импульсов через лимбические области, возможно увеличивая степень внутренней обратной связи с этими зонами (или между этими зонами и неокортексом [новой корой]), затмевая тем самым входные сенсорные сигналы. Также возможно и изменение пропускания сигналов, связанных с тета-ритмом, через заднюю часть поясной извилины (311).
Сведение всего это вместе в некоторое подобие согласованной теории может быть преждевременным, но, в любом случае, я собираюсь проделать это. Я считаю (все это может перемениться с приходом новой информации), что пониженный уровень сенсорных данных (из-за торможения "сверну вниз"), ослабленное кодирование промежуточной памятью, сопряженное с расширением активности коры задней части поясной извилины и $retrosplenial cortex$, - все это приводит к усилению устойчивости петли обратной связи. В этой петле случайный "шум", индивидуальные различия в строении и "разводке" лобных долей, содержимое промежуточной памяти и влияние электромагнитных полей (332) в комбинации производят глубоко анормальнфе нейропаттерны.
Интересный побочный эффект всего этого - усиление способности распознавать запахи. Эта способность формально не изучалась, и на самом деле все может происходить только в голове, однако обонятельные сигналы обрабатываются мозгом немного по-другому, чем остальные, и они могут не попадать под тормозящие импульсы, ослабляющие другие сенсорнные сигналы. И если это так, то петля обратной связи может действительно служить усилителем обонятельных данных, постоянно прибавляя одни и теже маленькие сигналы друг к другу.
9.2.6 Вклад непрямой активности
Многие из эффектов ДХМ несомненно возникают из-за его непрямой активности в других медиаторных системах. Например, он может ненапрямую увеличить 5HT [серотонин] активность, особенно в 5HT1A-рецепторе. Это может объяснить способность ДХМ изменять настроение. Еще один пример - дофаминергическая активность; ДХМ довольно сильно влияет на увеличение дофаминовой активности (и из-за активации сигма-рецепторов, и из-за предотвращения обратного захвата дофамина в PCP2-сайтах) (72,76). Так же показано, что блокада NMDA-рецепторов усиливает дофаминовую активность, а также активность других медиаторных систем (100).
9.2.7 Флэнджер
Одним из выдающихся эффектов ДХМ - флэнджер входящих сенсорных сигналов. В разной мере это проявляется у различных наркотиков. Ниже я изложу свою гипотезу на эту тему. Заметим, в частности, связь флэнджера с торчем [stoning] и опьянением [buzzing] - в некотором роде флэнджер - это более глубокая степень торча. Некоторые отметили флэнджер или строб-эффект после выкуривания большого количества каннабиса, потребители закиси азота знакомы с флэнджером звуков. Этот эффект может производить и алкоголь.
У всех этих веществ одна общая черта - они способны тормозить долговременную потенциацию в гиппокампе. Есть предположения, что существует несколько наборов сигналов, проходящих через лимбические области. Сигналы в фазе с тета-ритмом и остальные проходят по-разному. Вероятно один набор сигналов сильнее вовлекаются в процесс запоминания, а остальные - сильнее в проведение сенсорных данных. Или, вероятно, что все наркотики, тормозящие долговременную потенциацию, активизируют кору задней части поясной извилины. В обоих случаях, прохождение сигналов в одной фазе c тета-ритммм может быть нарушено, что приведет к пульсирующему восприятию в темпе тета-ритма.
Есть и другая теория - нейросети в мозгу повторно обрабатывают одни и те же данные, пока не сформируется устойчивая кофигурация, а ДХМ (и другие наркотики) ослабляют и замедляют этот процесс. Когда одни нейросети заторможены, а другие работают с нормальной скоростью, характеристические частоты двух наборов сигналов будут различаться, приводя к "биению частоты" почти тем же самым путем, как и два очень близких звука приводят к биению частоты (краткая демонстрация этого - послушайте как настраивают гитару на слух).
9.2.8 Гиперабстракция
Другой интересный эффект ДХМ - эго способность вызывать своеобразные когнитивные нарушения, которые я объединяю под термином "гиперабстракция". Два примера:
- Мем (meme) - "частичка" или "вирус" мысли - идея, которая в своем роде содержится сама в себе и распространяется как вирус. Например, идея гражданских свобод - это мем, который в некоторый момент обрел существование, быстро распространился, а сейчас стал интегрированной частью нашего сознания. Один потребитель ДХМ во время трипа внезапно осознал (или выдумал) "самоосуществляющийся, самотворный мем", который был концепцией мема, чье отождествление, понимание создает и пробуждает его. Кажется, что этот мем был безвременным в смысле, что он должен был всегда существовать, или не мог прийдти в голову, поскольку он не может быть выведен ни из чего, кроме самого себя (разумеется, мозг не работает по правилам логики, но ... вы поняли фишку).
- Другой потребитель описал мысли о сходящихся бесконечных суммах (т.е. 1/2 + 1/4 +1/8 и т.д., что в сумме дает 1). Хотя их можно складывать до бесконечности, проще абстрагировать процесс и получить ответ таким путем. Этот человек представил бесконечные последовательности абстракций, а затем - абстагировал эти бесконечные последовательности, чтобы выйти на новый уровень абстракции.
Многие вызванные ДХМ мысли включают в себя то, что иногда называется "Странными Циклами" в логике. Точно так же как и самопротиворечивое утверждение "это утверждение ложно", некоторые из них не могут быть воплощены в логической форме; другие - могут, но только в качестве гипотез. Мыслить на таком уровне абстракции очень нелегко (если вы конечно не Курт Гедль).
Некоторые люди, описавшие от первого лица случаи психоза и шизофрении, отметили увеличение абстрактных способов мышления (на ум приходит книга Дзен и Искусство Ухода За Мотоциклом Роберта М. Персига). Это может быть, разумеется, и полной чушью - эти абстрактные мыслеформы просто усиливающееся безумие (а в этом случае трудносопоставимы с конкретными идеями). С другой стороны, может быть, что нечто типа шизофрении и психотического состояния связано с "размазыванием" между уровнями абстракции. При достаточном "размазывании", мысль, немогущая быть представленной в деталях на конкретном уровне абстракции, сможет стать представимой в сознании.
Итак, ДХМ может вызвать нечто типа временного "размазывания" этих уровней абстракции. Происходит ли это благодаря NMDA или сигма-активности, я не знаю, хотя подозреваю последнюю, поскольку другие NMDA-антагонисты не имеют тенденции производить такие изменения в образе мышления.
9.2.9 Бред и проблемы с памятью
Как говорилось выше, сигма-активность может модулировать холинергические рецепторы в мозге (97), что приводит к временному снижению их функционирования аналогично (но гораздо более безвредному) действию антихолинергиков - атропина, скополамина, циклизина (Marezine) и т.д. Известно, что холинергическая активность важна для запоминания, а многие ноотропы (Умные наркотики - "Smart Drugs") усиливают холинергическую функцию. Сигма-активность может на время понизить эффективность в некоторых холинергических рецепторах, что приводит к нарушению памяти и мыслительного процесса. Некоторые люди фактически отмечали, что ДХМ создает ощущение временной тупости (Тупые наркотики, а?), хотя это происходит не со всеми.
Многие биогенные аминовые системы, видимо, играют управляющую роль, и многие исследователи считают, что эти управляющие системы работают как "пусковые кнопки". Например, одна из теорий ЛСД говорит, что он изменяет "коэффициент усиления" распознающих нейросетей (вероятно уменьшая торможение, хотя и иными путями, чем постулировано для диссоциативов). Как следствие, случайный шум (необходимый для работы любой нейросети) становится гораздо сильнее входящей сенсорной информации. Сенсорное распознавание становится все менее и менее точным - следовательно - галлюцинации.
Эффекты ЛСД почти наверняка более сложны, чем описано, но идея "пусковых кнопок" может быть справедливой и в этом случае. Если это так, то холинергические системы могут быть "пусковыми кнопками" для когнитивных сетей в той же мере, какими 5HT2A/5HT2C-системы являются для сенсорных нейросетей распознавания. Бред может быть просто когнитивным эквивалентом галлюцинаций. Или другими словами, разница между ощущением себя "похожим на цветок" и мыслями "Я и ЕСТЬ цветок" может быть ответом на вопрос какая нейросеть поражена.
Проблемы с памятью в некотором роде происходят из-за NMDA-блокады, хотя некоторые потребители кетамина отмечали, что ДХМ сильнее влияет на память, чем кетамин. Возможно, что в добавок к наведению бредовых мыслей, ухудшение холинергических функций может быть виновно в проблемах с памятью. Это разумеется согласуется с эффектом галлюциногенных (delusional) антихолинергиков [как, например, тарен].
Между прочим, антихолинергики тоже воздействуют на ацетилхолиновые рецепторы, которые управляют работой сердца и дыханием (эти рецепторы видимо не управляются посредством сигма-активности). Рекреационное использование антихолинергиков может быть чрезвычайно опасным, приводя к коллапсу дыхания и сердечной недостаточности.
9.3 Почему у ДХМ существуют плато?
9.3.1 Плато 1-3: Совокупность рецепторов
На рисунке изображен вероятный эффект совокупности рецепторов. Заметьте - это качественный рисунок, а не количественный;
настоящие уровени насыщения различных рецепторов для данной дозы ДХМ все еще практически неизвестны. Кривые означают уровни
насыщения в PCP2, сигма1 и сайте связывания в открытом канале NMDA, полное насыщение соответствующего
рецептора обозначено "уплощением" кривой. Ось Х -концентрация ДХМ; Y - процент насыщения рецепторов.
Между прочим, если вы знакомы с вещами подобного рода, вам может быть удобнее пользоваться координатами Lineweaver-Burk [что это - я не знаю], но для непосвященных это может оказаться смущающим.
Из-за повышенного сродства к PCP2, сигма1 и NMDA-рецепторам соответственно (сигма2 не представлен), малая доза будет оказывать пропорционально больший эффект на PCP2-рецепторы, в то время как увеличение дозы приведет к насыщению этих рецепторов. Прием большего количества ДХМ не изменит уровень насыщения PCP2 существенным образом, а окажет влияние на другие рецепторы.
Более того, более слабые эффекты воздействия на PCP2-рецепторы может быть почти полностью сглажены эффектами сигма1 и NMDA-рецепторов (различные уровни насыщения показывают это). Это совершенно разумно, поскольку и сигма1 и NMDA-активность выявляют довольно сильное изменение поведения, последний в большей степени, чем первый.
Итак, первое плато может соответствовать доминированию PCP2-активности с некоторой сигма-активностью и небольшим эффектом NMDA-блокады; второе плато - сигма и некоторым NMDA-эффектам; третье - глубокой NMDA-блокаде.
Это, несомненно, упрощение, и не принимает во внимание индивидуальные отклонения в рецепторах. Некоторые люди могут иметь генетические варианты рецепторов, к которым ДХМ имеет более сильное (или слабое) сродство. Человек с PCP2-рецепторами, сильно связывающимися с ДХМ, может наслаждаться стимулирующими эффектами гораздо больше других.
К тому же, ионные каналы и сигма2-рецепторы в рисунке не указаны. Однако они несомненно вносят свой вклад, и некоторые люди утверждают, что есть плато и между первыми тремя. Они могут быть неярко выраженными и незаметными для большинства.
9.3.2 Четвертое Плато: Отключение чувств
А что происходит на четвертом плато? Ну, вот еще один маленький рисуночек, который, надеюсь, прояснит все (ага, точно). Это рисунок нейросети; точки - нейроны, линии - их соединения. Как и все в мозгу, этоа нейросеть сильно связана. Проценты - количество оставшихся функциональных связей.
Когда достаточное число NMDA-рецепторов блокировано, нейрон может потерять большую часть входящей информации от другого нейрона и соединение эффективно нарушается. В начале это не причиняет неудобств, поскольку нейроны и связи нейронов еще достаточно плотны и другие нейроны могут взять на себя дополнительную нагрузку (хотя результатом может стать более медленная и менее аккуратная работа нейросети). В некоторый момент, достаточная связанность теряется и нейросеть уже больше не может функционировать.
Сравните это с диссоциацией четвертого плато. На некотором уровне, некие части мозга (вероятно гиппокамп) теряет способность к функцинированию настолько, что больше не может работать как единый блок. Сенсорный процессинг останавливается, и необработанная входящая сенсорная информация не может быть преобразована в соответствующе проанализированный (parsed) выходной поток. Сознание остается без реальной информации; вместо этого образуются хаотические нестабильные мыслеформы (паттерны). Следовательно - диссоциативная анестезия.
С другой стороны, может случиться так, что порог четвертого плато достигается, когда сенсорная информация тормозится достаточно для того, чтобы нейросигналы в лимбических областях отражались назад (feed back) на вход без существенного подавления со стороны входящей сенсорнной информации. Эта петля обратной связи продолжает возмущать нейропаттерны до тех пор, пока они не становятся оторванными от состояния человека или его воспоминаний.
9.4 Почему это так сложно?
ДХМ сам по себе очень сложное вещество; большинство наркотиков связыватся с одним или двумя рецепторами (покрайней мере с одним классом рецепторов). Рекреационное злоупотребление им, известное уже догие годы, еще не изучалось должным образом, он может воздействовать по-разному на разных людей. Рецепторы и сайты связывания, на которые он воздействует - сигма, NMDA и PCP2 - недавние открытия. Все это приводит к сложному и полохо понимаемому наркотику.
Кроме того, мозг - это сложная система, и, по большому счету, мы ничего не понимаем в его работе. Основы нейропередачи (нейротрансмиссии) вроде бы поняты, но остается много вопросов. Никто не знает почему так много нейромедиатров или подтипов рецепторов. Еще хорошо не изучены системы вторичных посредников большинства рецепторов. Большинство событий внутри нейронов происходит путем изменений в экспрессии генов, а это отдельная тема, о которой мало известно.
Повторю часто цитируемые (и правильные) слова: если бы наш мозг можно было бы просто понять, мы бы были такими примитивными, что не смогли бы его понять. Я верю, что в конце концов мы узнаем как работает мозг, но это случится не при моей жизни. До тех пор мы просто пытаемся догадаться, шаря в темноте.
9.5 Фармакокинетика: Как метаболизируется ДХМ
ДХМ, как соль гидробромида, быстро абсорбируется из желудочно-кишечного тракта; в течении 30 минут он может войти в кроветок (2,3). Соединение полистирекса предназначено для продолжительной абсорбции, и его полный переход в кровь может занять 6-8 часов.
ДХМ метаболизируется двумя путями, оба из них приводят к одному - 3-гидроксиморфинану (3ГМ, 3HM). Первый путь исходит из ДХМ к ДХО (декстрорфан), а затем к 3HM; второй из ДХМ к 3-метоксиморфинану (3ММ), а затем к 3HM. Большая часть ДХМ (до 90%) метаболизуется через ДХО у нормальных индивидуумов.
ДХМ преобразуется в ДХО печеночным ферментом, называемым цитохром P450-2D6 (дебрисохин 4-гидроксилаза, debrisoquine 4-hydroxylase). Примерно у 7% кавказцев и 0,5% азиатов версия этого фермента работает крайне (до 70 раз медленнее) неэффективно и не может метаболизировать ДХМ в ДХО должным образом (10). От 0.5 до 2% популяции имеют многочисленные копии гена P450, что приводит к очень быстрому метаболизму ДХМ в ДХО (155). После превращения в ДХО, ферменты P450-3A4 и P450-3A5 перобразовывают ДХО в 3-гидроксиморфинан(77).
Другой путь ведет сначала к 3-метоксиморфинану (посредством ферментов P450-3A4 и P450-3A5), а уже затем к 3-гидроксиморфинану. Большинство людей не метаболизуют основное количество ДХМ таким образом, хотя люди с недостатком нормального P450-2D6 будут превращать существенную часть ДХМ в 3ММ. А 3ММ вероятно не психоактивен, что означает - 5-10% людей с недостатком фермента 2D6 ощутят меньший эффект от ДХМ (или более точно, не испытают эффектов ДХО).
P450-2D6 работает, убирая метокси группу и заменяя ее гидрооксилом (OH) (или точнее, отрезая метил от кислорода); этот процесс называется О-деметиляцией. P450-3A4 и 3A5 заменяют метильную группу водородом (H); это процесс N-деметиляции. Положение метильной и метокси групп показано на рисунке.
9.5.1 Факторы, влияющие на метаболизм ДХМ
Как говорилось ранее, у некоторых людей отсутствует нормальный фермент P450-2D6. У остальной популяции этот фермент может быть ингибирован несколькими факторами. Многие лекарства ингибируют P450-2D6, особенно флюокситин (ProzacTM). Неполный список лекарств, ингибирующих фермент P450-2D6, дан в Разделе 15.1.
Сам по себе ДХМ, разумеется, будет конкурировать с другими веществами за P450-2D6 и, что важно, это будет делать и 3-метоксиморфинан (3ММ) (17). Фактически, 3ММ имеет большее сродство к ферменту P450-2D6, чем ДХМ. Это может объяснить тот факт, что вторая доза ДХМ "в догонку" призводит эффект отличный от первой. Конкуренция за фермент P450-2D6 уменьшит количество ДХМ, превращающегося в ДХО у второй дозы.
Этот график получен путем компьютерной симуляции метаболизма ДХМ:
Первая пара графиков представляет метаболизм ДХМ у нормального индивидуума (слева) и индивидуума без нормального фермента P450-2D6 (справа). Заметьте быстрое и почти полное превращение ДХМ в ДХО у нормального индивидуума, по сравнению с менее эффективным и более медленным превращением у человека с недостатком P450-2D6.
Вторая пара демонстрирует возможные результаты приема дополнительных доз ("в догонку"). Оба графика соответствуют нормальному варианту фермента P450-2D6. Отмечу, что вторая доза ДХМ не превращается в ДХО также быстро (поэтому меньший наклон). Правый график показыавет многочисленные дозы, и выгибание вверх метаболитической кривой ДХМ становится все более явным с каждой дозой.
Между прочим, это все качественные симуляции, а не количественные. Я пытался придерживаться известных значений КМ и VMAX для подходящих реакций [?], но симуляции были дискретным процессом (если честно, мои познания в уравнениях довольно слабы и требуют от меня титанических усилий). Я сравнил мои результаты с обнаруженными скудными данными, сравнения оказались разумными, но, опять же, я могу быть абсолютно неправым. Цель этих графиков - продемонстрировать относительные эффекты изменений в ферментной активности (из-за генетических изменений и конкурентного ингибирования 3-метоксиморфинаном), надеюсь, что этого оказалось достаточно.
У меня нет информации о том, что случается с самим 3-гидроксморфинаном. Он может выделяться напрямую почками, или подлежит дальнейшему метаболизму.

| 
| 
|